Triose phosphate/phosphate translocator (TPT)
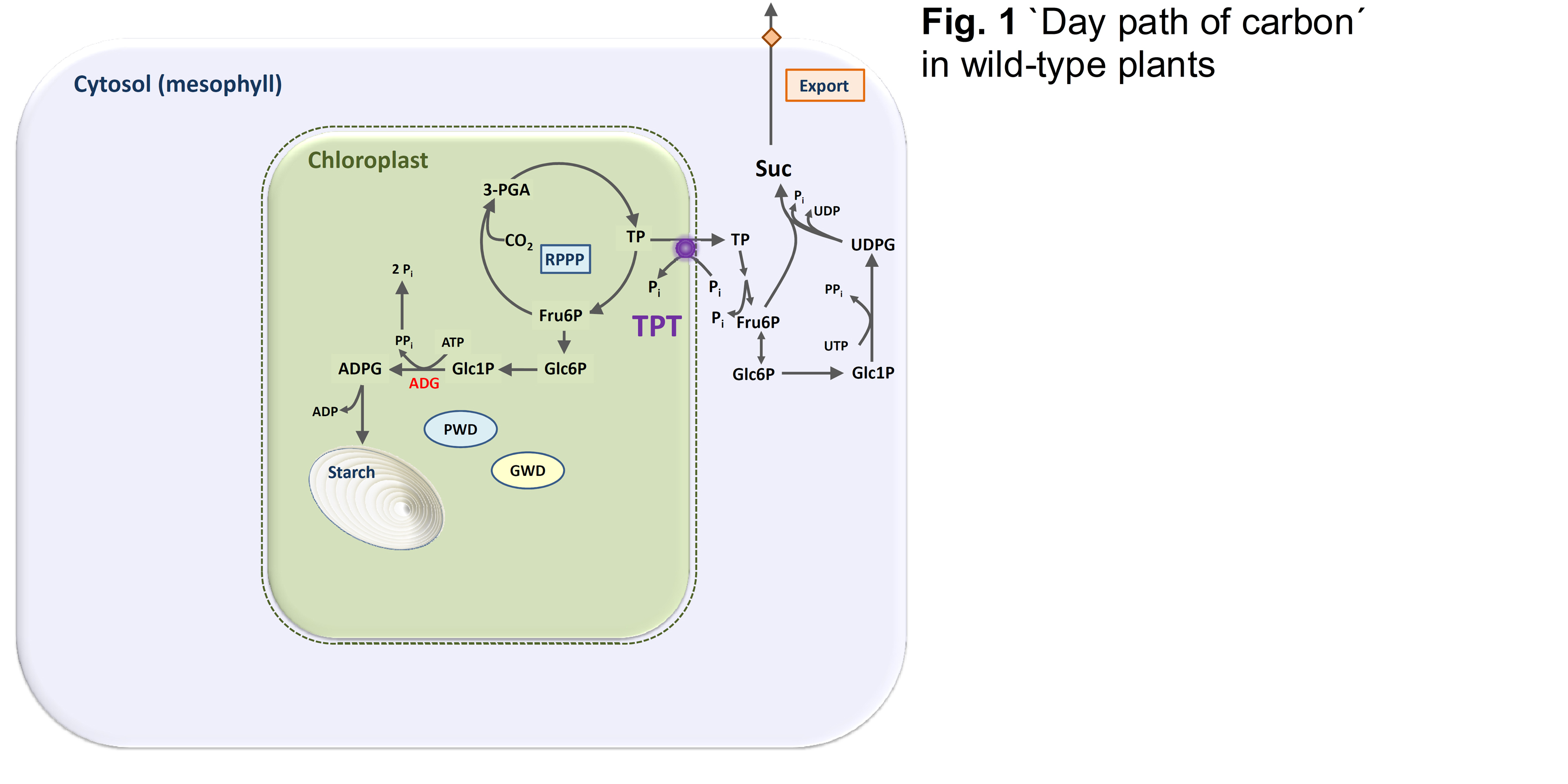
The triose phosphate/phosphate translocator (TPT) mediates the export of photoassimilates from the chloroplasts in form of triose phosphates in the light. This `day path of carbon´ supports sucrose biosynthesis in the cytosol for export to heterotrophic tissues and organs (Fig. 1).
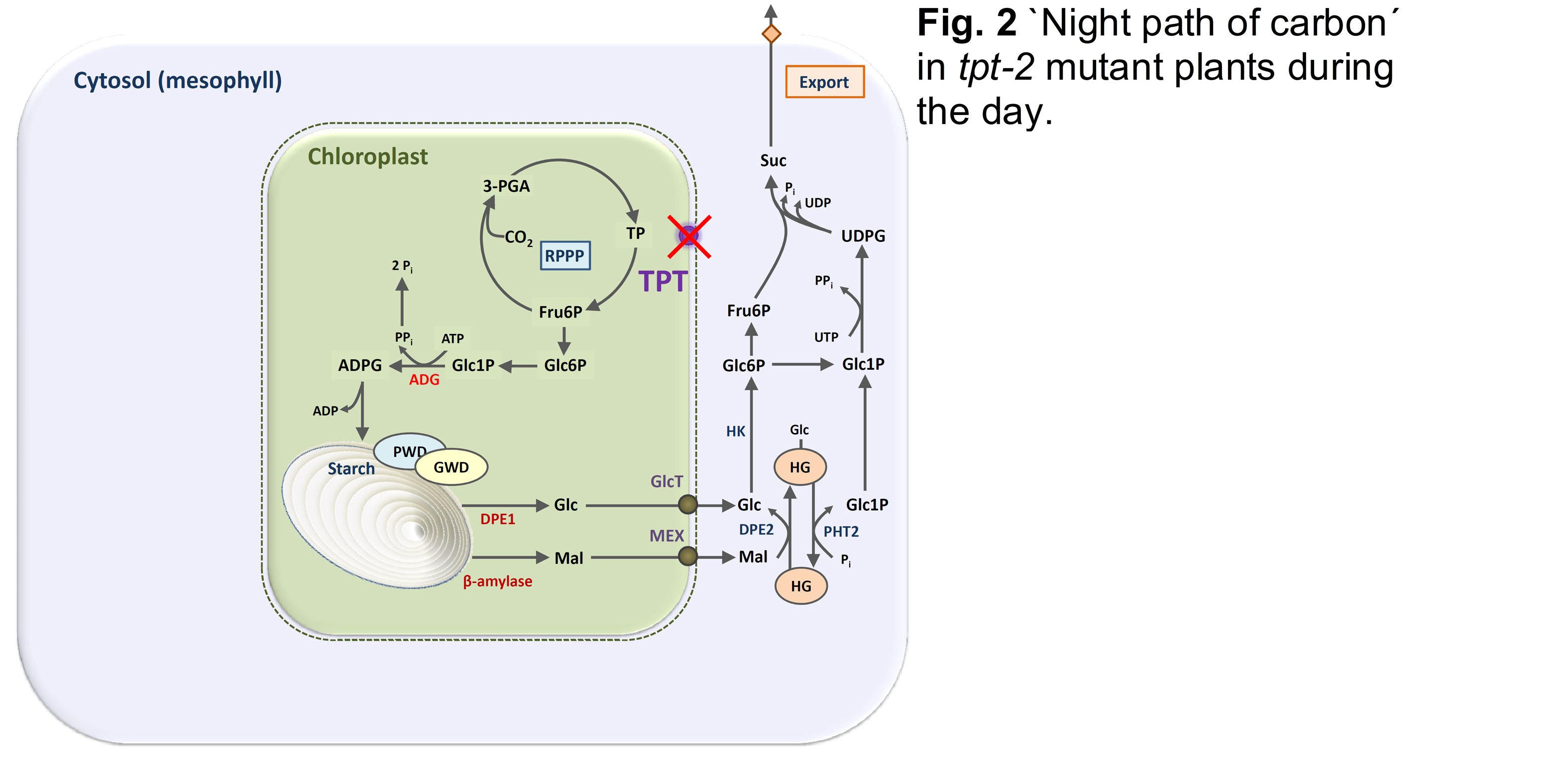
The loss of TPT function can be compensated for by an enhanced `night path of carbon´, i.e., the mobilisation of transitory starch in the light and dark (Fig. 2; Häusler et al. 1998; Schneider et al., 2002).
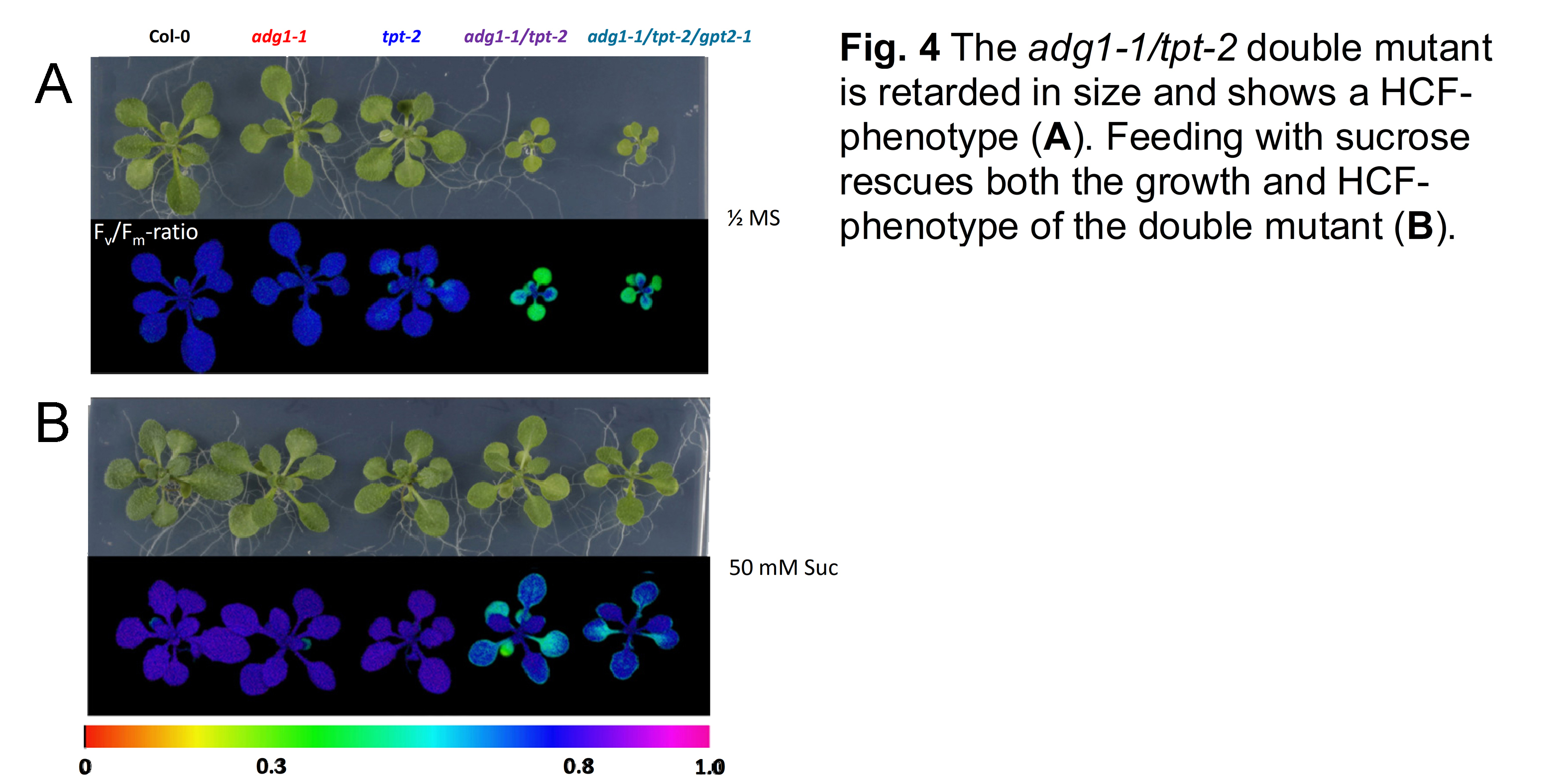
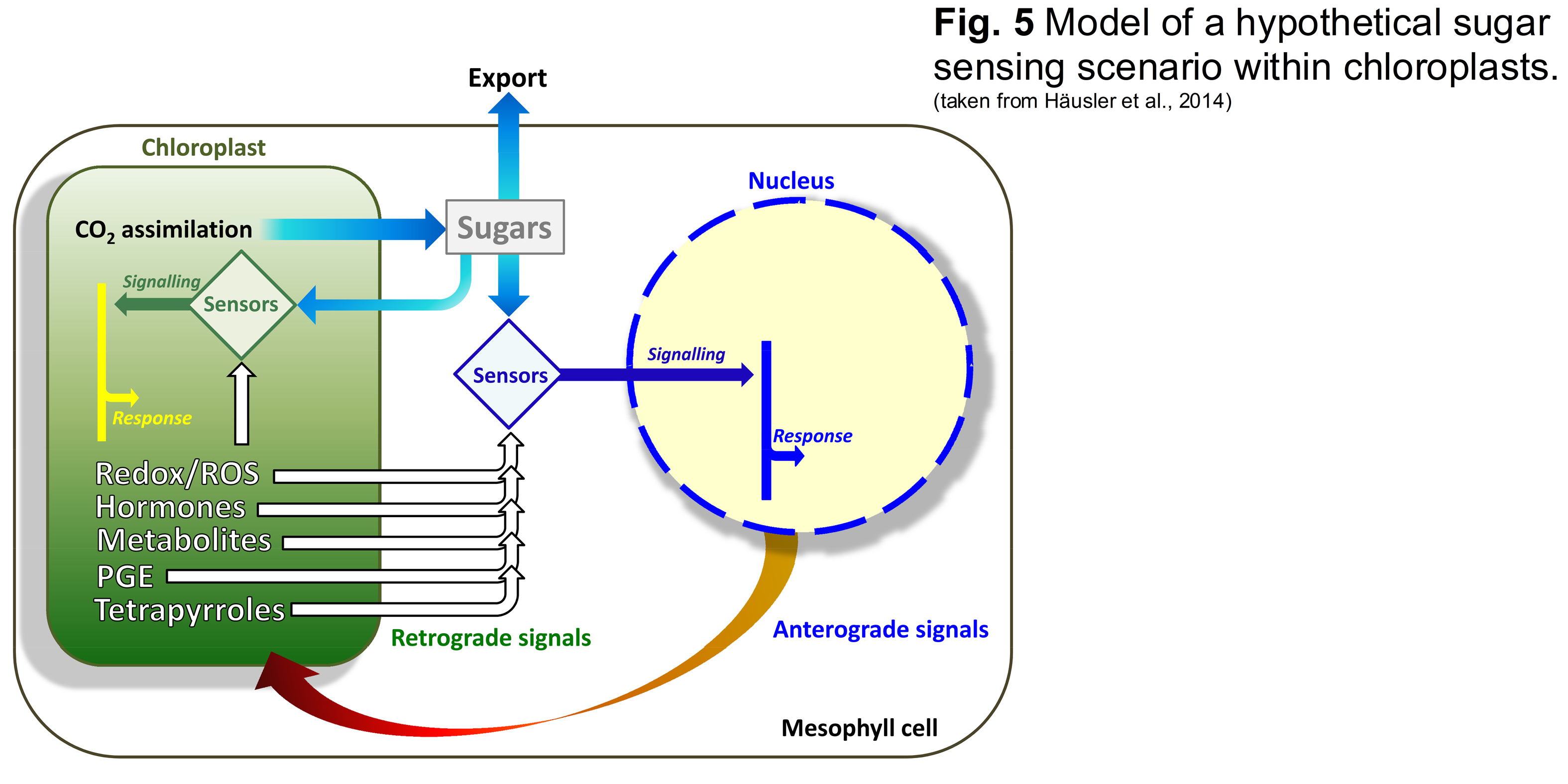
Only if starch biosynthesis is inhibited in combination with a knockout of the TPT, e.g., in the adg1-1/tpt-1 (Schneider et al., 2002) or adg1-1/tpt-2 double mutants (Schmitz et al., 2012; Fig. 3) the plants are severely retarded in size and show a high chlorophyll fluorescence (HCF) phenotype in the dark-adapted state (Häusler et al., 2009; Fig. 4). The rescue of both phenotypes by feeding sugars (Heinrichs et al., 2012; Schmitz et al., 2012) as well as by growth of the plants at low light intensities gave rise to the idea that the double mutants might be impaired in retrograde signaling, responsible for the synchronised expression of plastome- and nuclear-encoded photosynthesis genes (Schmitz et al., 2014; Gläßer et al., 2014). The rescuing response of sugar feeding has been interpreted as altered sugar sensing within the chloroplasts (Häusler et al., 2014; Fig. 5). A hypothesis awaited to be addressed experimentally.
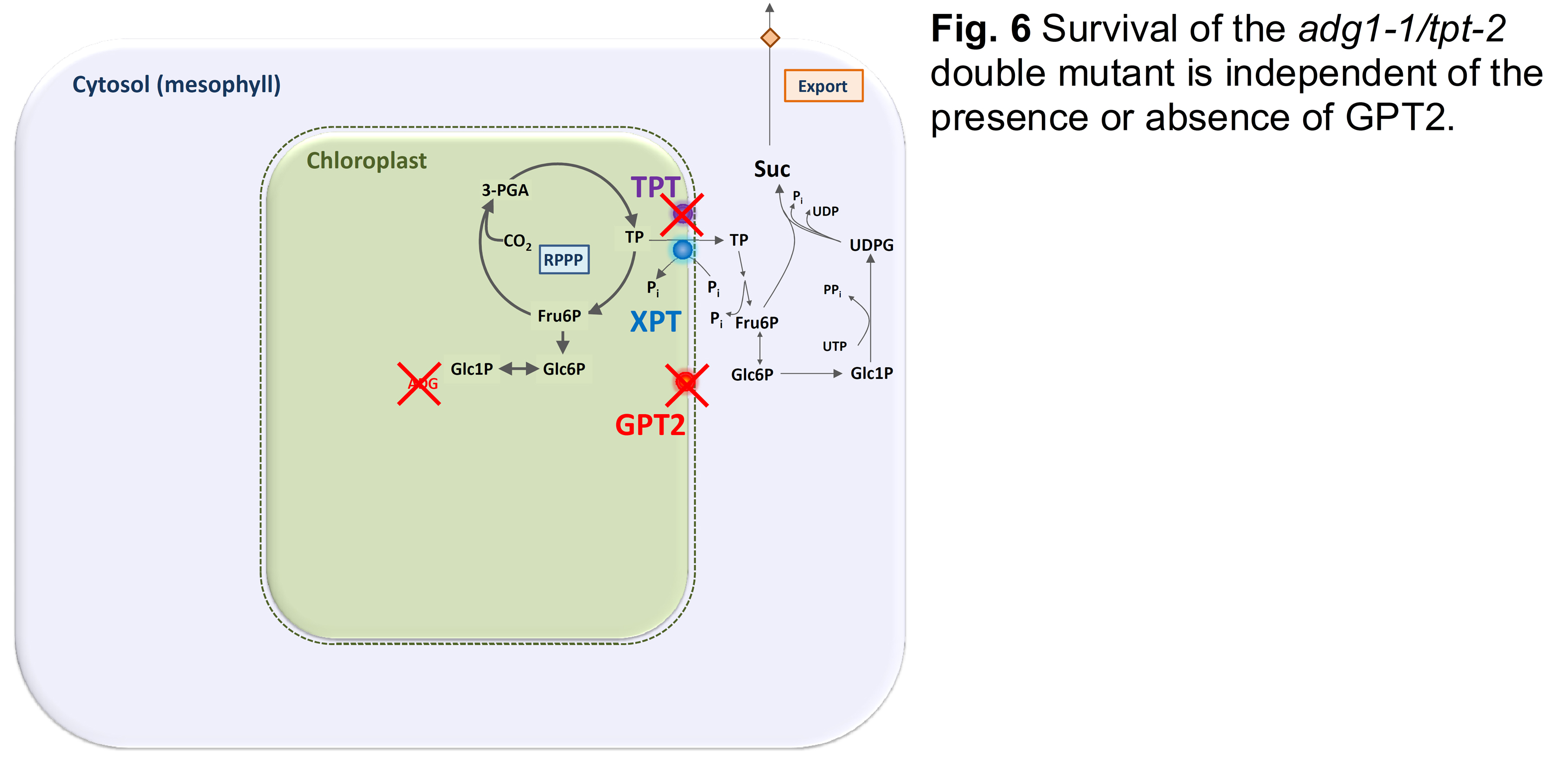
Further analyses revealed that the viability of adg1-1/tpt-2, in particular its rescue by feeding sugars is independent of GPT2 (Fig. 6) as adg1-1/tpt-2/gpt2-1 triple mutants could not be distinguished from the adg1-1/tpt-2 double mutant (Schmitz et al., 2012; see Fig. 4).
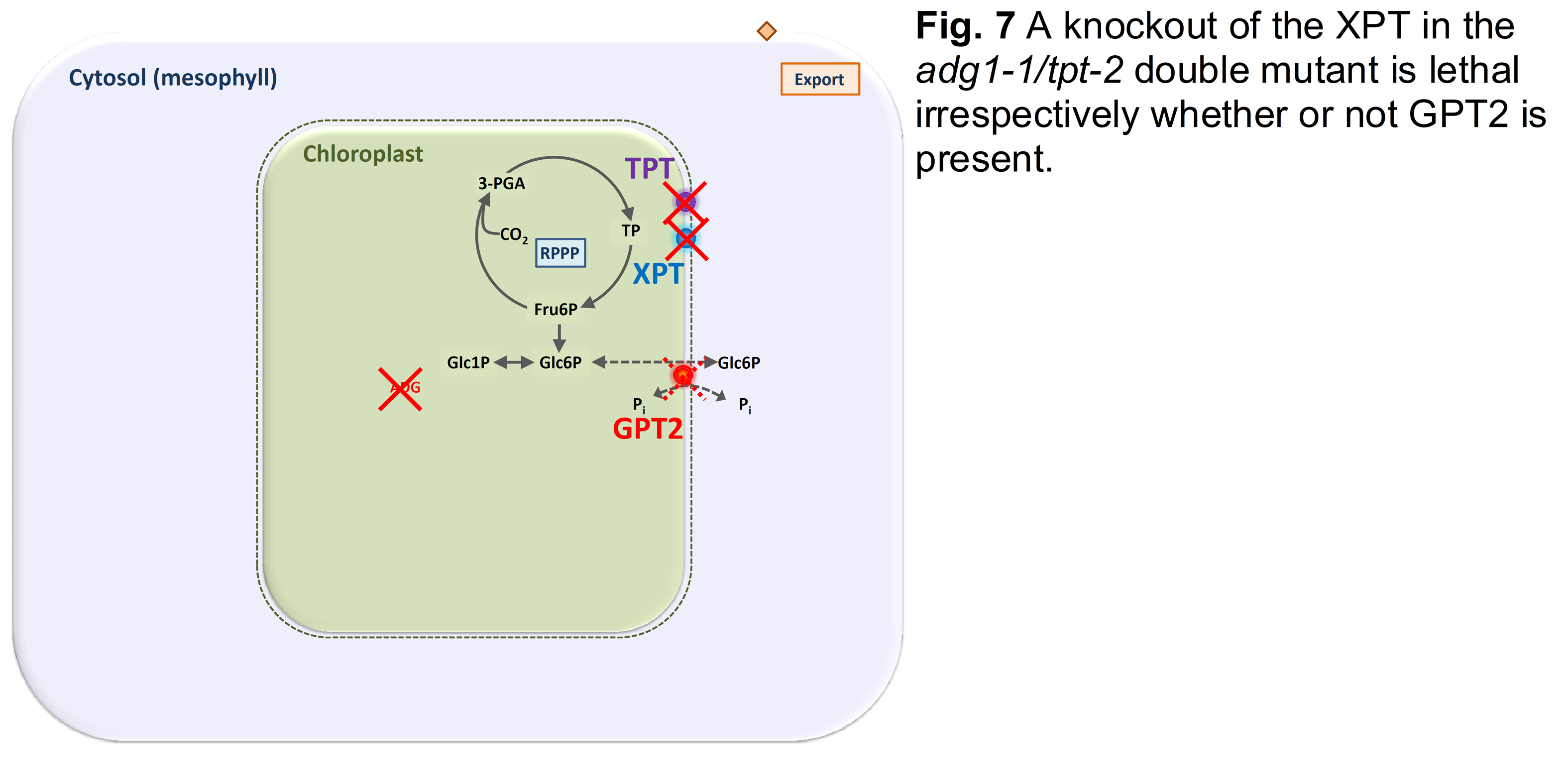
Only if the XPT is simultaneously knocked out in the background of the adg1-1/tpt-2 double mutant, the resulting triple mutant is lethal (Hilgers et al., 2018; Fig. 7).
(back to Rainer E. Häusler)